

The medicinal fungus Cordyceps militaris: Research and development
Abstract Ophiocordyceps sinensis (syn. Cordyceps sinensis) is a highly valued medicinal fungus. This entomopath-ogen has a limited distribution, has been overharvested in the wild, and its stromata have not been artificially cultivated. Another entomopathogenic fungus, Cordyceps militaris (commonly known as orange caterpillar fungus), has chemical capacities similar to those of O. sinensis, but unlike O. sinensis, its stromata can be easily cultivated. Consequently, C. militaris is being studied as an alternative to O. sinensis, and the large-scale production of stromata is receiving substantial attention. Significant research has been conducted on the genetic resources, nutritional and environmental.

B. Shrestha
Green Energy Mission/Nepal,
Ghatte Kulo, Anam Nagar,
Kathmandu, P.O. Box 10647, Nepal
W.Zhang
Guangdong Institute of Microbiology,
Guangzhou 510070, China
Y. Zhang
School of Life Sciences, Shanxi University,
Taiyuan 030006, China
X.Liu (*)
State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences,
No.3, 1st West Beichen Road, Chaoyang District,
Beijing 100101, China e-mail: liuxz@im.ac.cn
Present Address:
B. Shrestha
Mushroom Research Division, National Institute of Horticultural and Herbal Science, Rural Development Administration,
Suwon 441-707, Korea
requirements, mating behavior, and biochemical and pharmacological properties of C. militaris. The complete genome of C. militaris has recently been sequenced. This fungus has been the subject of many reviews, but few have focused on its biology. The current paper reviews the biological aspects of the fungus including host range, mating system, cytology and genetics, insect- and non-insect nutritional requirements, environmental influence on stroma development, and commercial development.
Keywords Cultivation. Entomopathogenic fungus. Mating system. Pharmaceutical application
Historical review
Medicinal fungi have long been an important part of human culture and civilization, and species in the genus Cordyceps are especially valued (McKenna et al. 2002). Most Cordyceps species s.l. are parasites of insects or other arthropods (Kobayasi 1941, 1982). Ophiocordyceps sinensis (Berk.) G.H. Sung et al. (syn. Cordyceps sinensis (Berk.) Sacc.), for example, is a parasite of caterpillars (Wang and Yao 2011) and is naturally distributed in the Tibetan Plateau of China and surrounding high-altitude grasslands of Nepal, Bhutan, and India (Shrestha et al. 2010; Zhang et al. 2012b). As a highly valued medicinal fungus, O. sinensis has a long history of being collected and traded (Jones 1997; Halpern 1999). Recent studies have shown that the natural populations of O. sinensis are decreasing because of overcollection (Li et al. 2006c; Stone 2008; Zhang et al. 2012b). Because of the scarcity and high price of wild O. sinensis specimens, many counterfeit materials including cultured products are sold in the market, resulting in the
Many Cordyceps species are morphologically or otherwise similar to C. militaris, and these include C. cardinalis G.H. Sung & Spatafora, C. kyusyuensis A. Kawam., C. pseudomilitaris Hywel-Jones & Sivichai, C. rosea Kobayasi & Shimizu, C. roseostromata Kobayasi & Shimizu, C. washingtonensis Mains, and others (Sung and Spatafora 2004; Sung et al. 2007; Wang et al. 2008). Although distinguishing C. kyusyuensis from C. militaris is difficult based on morphology and cultural characteristics (unpublished work), the larvae parasitized by C. kyusyuensis tend to be larger and located deeper in soil than larvae parasitized by C. militaris (Kawamura 1955; Sung 1996; see Table 1). C. kyusyuensis, originally described from Japan on larvae of the lepidopteran Clanis bilineata (family Sphingidae) (Kawamura 1955), has been reported only from China (Kobayasi 1981; Guo and Li 2000) and Korea (Sung 1996), besides Japan. According to Wang et al. (2008), C. kyusyuensis is not a different species, rather a synonym of C. militaris.
Mating system
Only a few cytological studies have been conducted on C. militaris. Earlier studies showed that C. militaris has only two chromosomes (Varitchak 1927; Jenkins 1934), while a recent study reported seven chromosomes (Wang et al. 2010c). Despite the scarcity of cytological studies, the mating system of C. militaris has been studied in many instances at both cultural and molecular levels. Inoculations of two single-ascospore strains together or on opposite sides on a fruiting medium plate are the two major methods for genetic analysis of mating system in ascomycetous fungi, including Cordyceps spp. (Harris 2001). Gao et al. (2000b) reported that single-ascospore strains of C. militaris were unable to produce stromata when growing alone on media. Later, Sung and Shrestha (2002) and Shrestha et al. (2004a) showed that single-ascospore strains of C. militaris produced perithecial stromata when inoculated in combination with a compatible isolate, but produced only deformed stromata without perithecia when inoculated singly. Further, it was shown that primordia developed in the meeting line between two opposite mating-type strains that were inoculated on opposite sides of an agar plate (Shrestha et al. 2004a). These results show that C. militaris is a bipolar heterothallic fungus. The mating type character was found to be stable for as many as 10 generations of subcultures of the original strains (Shrestha et al. 2004a; Sung et al. 2006a). Each single ascospore strain was found to be hermaphroditic but self-incompatible (Shrestha et al. 2004a). In agreement with these studies, Liang et al. (2005) observed that, when growing alone, most of the single-ascospore strains could not produce well-developed stromata or produced only abnormal ones. Gao
and Zheng et al. (2011a) also recently confirmed bipolar heterothallism in C. militaris. A molecular study has demonstrated that C. militaris possesses opposite mating-type idiomorphs, MAT1-1 and MAT1-2 (Yokoyama et al. 2006), or two mating type genes, MAT-a and MAT-HMG (Wang et al. 2010a). Zheng et al. (2011a) recently identified the MAT1-1 mating type locus that included MAT1-1-1 and MAT 1-1-2 genes.
Despite obvious heterothallism, homothallism has been occasionally observed in C. militaris (Sato and Shimazu 2002a; Shrestha et al. 2004a; Liang et al. 2005; Wen et al. 2009; Zheng et al. 2011a). Homothallism in C. militaris could be due to the presence of (1) both genotypes (MAT1-1/2) in the chromosomes, (2) a disomic (diploid) condition with an extra chromosome of the opposite mating type locus, or (3) a heterokaryotic condition with more than one nucleus with opposite mating type loci (Shrestha et al. 2004a; Wen et al. 2009). Intraspecific high genetic variation seems common in C. militaris and may be associated with mating-system instability (Wen et al. 2009). Recently, het-erokaryosis and parasexuality were confirmed in singlespore strains of C. militaris by RAPD analysis (Li et al. 2007a). Mating-type switching can also be another possible reason for homothallism in C. militaris.
In contrast to mating-type loci in homothallism, mating-type loci in heterothallic fungi exist in separate spores. Mating compatibility in heterothallism can be controlled by only one factor (bipolar) or by two factors (tetrapolar). Basidiomycetous fungi are mostly heterothallic, while a majority of ascomycetous fungi studied are homothallic. Heterothallic ascomycetous fungi contain a single mating-type locus with two alternate alleles (bipolar) (Poggeler 2001), while basidiomycetes have evolved multiple mating-type loci (Kothe 2001). To determine mating compatibility in ascomycetes, a researcher must wait until fertile reproductive structures are formed. There is no specific indication in the somatic structure or the asexual reproduction structure in C. militaris strains that they have different abilities of producing fruiting-bodies (Gao et al. 2000b; Wu et al. 2000b). Liang (1990, 2001), however, distinguished fruiting-body formation from ‘Paecilomyces-type’ but not from ‘Acremonium-type’ strains. Li et al. (2006a) also reported that fruiting-bodies were developed from a ‘Lecanicillium-type’ strain but not from an isolate with more densely verticillate phialides. In another study, it was shown that fast-growing isolates produced fewer fruiting-bodies than slow-growing isolates (He et al. 2010).
Although C. militaris lacks distinctive phenotypic characteristics both in the wild and in culture, one obvious phenotypic characteristic in culture is the colony pigmentation that ranges from orange to yellowish-white on nutrient-rich agar media (Shrestha et al. 2006). Either kind of pigmentation was observed in F1 progeny strains when two.
parent strains with contrasting pigmentations (i.e, yellowish-white vs. orange) were crossed (Shrestha et al. 2005a). The F1 progenies either on back-crossing with the parents or sister-crossing among themselves showed that colony pigmentation was independent of or distantly linked with the mating-type locus (Shrestha et al. 2004a, 2005a).
The study of the mating system is more difficult with clavicipitaceous fungi than with many other ascomycetous fungi. Clavicipitaceous fungi are characterized by septate, filamentous ascospores. From a cytological perspective, each filamentous clavicipitaceous ascospore corresponds to a unicellular ascospore of other ascomycetes. Ascospore initials in C. militaris undergo asexual or somatic division seven times and are ultimately filamentous in shape and consist of 128 part-spores (Moore 1964; Ding et al. 1995; Hywel-Jones 2002).
Several characteristics of C. militaris ascospores make single-spore isolation difficult. Firstly, C. militaris asco-spores that consist of as many as 128 part-spores, also known as secondary spores, readily fragment into short filaments upon discharge from mature perithecia, rendering complete unfragmented ascospores difficult to find. Secondly, short filaments of the ascospores tend to overlap or attach end-to-end when released (Shrestha et al. 2005c), which often results in the isolation of multiple ascospores rather than a single one. Thirdly, ascospores tend to germinate quickly in agar media, and germinating hyphae can link the genomes of nearby ascospores (Liang et al. 2005; Shrestha et al. 2005c; Gao 2008). To avoid these problems, researchers should pay attention to ascospore length during isolation and should avoid isolating ascospores that appear abnormally long. Our experience is to leave the ascospores on the isolation medium for a few hours so that they swell but do not germinate. The swelling makes it easier to observe individual ascospore filaments. Although individual ascospore filaments can be isolated after some experience, it is not possible to isolate all the eight ascospores from the same ascus in a Cordyceps (s.l.) species. Isolation of individual part-spores is the final goal that can be achieved to establish a pure culture. Despite difficulty, it is hence urged , in future, to cultures from individual part-spores, probably attainable by the dilution method, in order to overcome the confusion of single-ascospore isolation.
Out-crossing has been successfully achieved for C. mil-itaris in culture. For example, Sung et al. (2006b) reported the formation of perithecial stromata from crossing between different specimens of C. militaris. Although it has been frequently studied, the mating system of C. militaris has been described as unknown in fungal textbooks (e.g., Webster and Weber 2007).
It is clear that mating compatibility not only produces fertile perithecia but also affects developmental and morphological patterns of stromata. Stromata produced by
successful mating are regular, club-shaped, or cylindrical, whereas those produced without mating are deformed and abnormal in size (Shrestha et al. 2004a; Liang et al. 2005). Usually, the cortex in the fertile clava of stromata becomes loose and spongy at maturity whereas, in those produced without mating, it remains hard and compact (unpublished data). The reason might be that substantial materials and energy are used for the development of perithecia in the case of mating, leaving the interstitial cortex tissue weak. Although the differences of biochemical composition between perithecial and non-perithecial stromata remain to be determined, Wen et al. (2005) showed that the amounts of medicinal components such as polysaccharide, adenosine, cordycepin, and cordycepic acid differ between sclerotium and stroma of C. militaris.
In addition to clarifying the life cycles of fungi, mating studies are also important for strain improvement (Kothe 2001; Poggeler 2001). In addition, the mating system also has evolutionary and taxonomic significance in Cordyceps. Although researchers continue to debate whether homothal-lism preceded heterothallism or vice versa (Geiser et al. 1998; Yun et al. 1999), it is generally assumed that the mating systems might have switched from one to the other type more than once in the course of evolution. Whether most Cordyceps species are homothallic or heterothallic remains unknown. A molecular study has shown that several Cordyceps species studied contain opposite mating-type idiomorphs, MAT1-1 and MAT1-2 (Yokoyama et al. 2006). A recent study of the mating system in O. sinensis has detected only the mating-type gene MAT1-2-1 of the idio-morph MAT1-2 (Zhang et al. 2011), suggesting that O. sinensis could be homothallic. The study of the mating system in O. sinensis is hindered by inadequate understanding of its biology, slow hyphal growth, difficulty in inducing stromata in culture, and the scarcity of genetic and genomic resources (Zhang et al. 2011).
In addition to increasing our understanding of fruiting-body formation, information about mating compatibility can also help with the identification of similar species, such as C. militaris and C. kyusyuensis, according to a “biological species concept”. It is important to understand the biological species concept of C. militaris in the context of its worldwide distribution and wide host range.
Nomenclature and status of the anamorph
Cordyceps militaris produces asexual spores (conidia) on the tips of little-differentiated phialides and eight filamentous sexual spores (ascospores) in an ascus (within a peri-thecium) during its sexual phase (Kobayasi 1941; Feng et al. 1990; Liang 1990; Ding et al. 1995). Naming of the conidial structure or anamorph began from the early phase of research on C. militaris, but the name has frequently changed.
Gams (1971) removed C. militaris anamorph from Paecilo-myces (now mainly Isaria) (Gams et al. 2005; Hodge et al. 2005) or Cephalosporium/Acremonium and transferred it to Verticillium sect. Prostrata. Zare and Gams (2001) later erected a new genus Lecanicillium to substitute for Verticil-Hum sect. Prostrata including the anamorph of C. militaris. It is a characteristic of this fungus that conidia can arise in the same strain either in chains or in heads which may be partially modified by different media. The phialides can either arise singly or in whorls again possibly dependent on the medium, but no mutation is required for such a change.
Finally, starting on 1 January 2013, the confusion over multiple naming of holomorphic fungi will be resolved through the recent revision of Article 59 of the International Code of Botanical Nomenclature (ICBN) [recently changed to the International Code of Nomenclature for algae, fungi, and plants (ICN)] that will validate only one name in a clade based on priority (Hawksworth 2011; Miller et al. 2011; Norvell 2011). The revision will help establish the concept of one fungus = one name (Hawksworth 2011). Different anamorph names associated with C. militaris can be stated as morphological stages, e.g., acremonium, lecanicillium, paecilomyces, verticillium, etc., with a lower case initial letter and normal non-italic type, following Cannon and Kirk (2000) and Hawksworth (2011).
Synonymous terms for stromata
A stroma is a compact, somatic structure or a cushion-like matrix of mainly ascomycetous fungi on which or in which spores or fruiting bodies are usually formed (Alexopoulos et al. 1996; Kirk et al. 2001). The stroma is generally distinguished from the synnema, which carries conidia or asexual spores at the tips of conidiogenous cells (phialides) or on indistinct hyphae. Stromata of C. militaris are clavate, clubshaped, or cylindrical with a lower sterile stipe and upper fertile clava, also known as the apical part or head, and are orange or yellow in color. In the wild, stromata are solitary to few or in some cases gregarious. The literature on the cultivation of Cordyceps spp., however, uses a variety of terms interchangeably with stroma, and these include fruiting-body (or fruitbody, fruit body, fruit-body), artificial stroma, perithecial stroma, sporophore, sexual sporophyte, sporocarp, ascostroma, and others. Among them, ‘fruit-body’, ‘fruit body’, and ‘fruiting-body’ are most commonly used and may be preferred to stroma for communication with farmers, businessmen, and consumers. Stroma and stromata, however, are the preferred terms for scientific reports. Different words have also been used to describe stromata produced in culture, and these include normal versus abnormal, complete versus incomplete, regular versus irregular, stable versus unstable, mature versus immature, perfect versus imperfect, and uniform versus deformed.
Cytology, genetics, and genomics
Only a few studies have been conducted on the cytology and genetics of C. militaris. Moore (1964) showed that the manner of somatic nuclear division is similar to that of cell division. Electrophoresis karyotype analysis showed that its chromosomal number is seven, and that chromosome size ranges between 2.0 and 5.7 Mb (Wang et al. 2010c). However, recent whole genome sequencing of C. militaris has shown that its total genome is exactly 32.2 Mb (Zheng et al. 2011a), which is smaller than that of two other entomopa-thogenic fungi, Metarhizium anisopliae (39.04 Mb) and M. acridum (38.05 Mb) (Gao et al. 2011). Conditions affecting the formation and regeneration of protoplasts from C. mili-taris have been extensively studied (Ma et al. 2008; Liu et al. 2009; Zhou and Luo 2009; Li et al. 2011), and mutants with superior traits, e.g., high production of cordycepin, polysaccharide, and fruiting-bodies, have been obtained by irradiation induction (Che et al. 2004; Zhou and Bian 2007; Zhou et al. 2009a; Li et al. 2011).
The study of fungal genetics requires an efficient transformation system. Zheng et al. (2011b) developed and optimized Agrobacterium tumefaciens-mediated transformation for C. militaris, which can facilitate the identification of functional genes. Expressed sequence tag (EST) analysis revealed different transcriptional patterns for C. militaris in mycelia growing in liquid culture or on solid rice medium, and in fruiting-bodies produced on rice medium or on silkworm pupae (Xiong et al. 2010). Further analysis showed that genes involved in cell metabolism, energy metabolism, and stress responses were upregulated during asexual development, and that genes associated with cell wall structures were upregulated during sexual development (Xiong et al. 2010). Many studies have described how conditions affect fruiting-body formation and metabolite production (e.g., cordycepin, polysaccharide) (Cui and Zhang 2011; He et al. 2011b), but the genetic basis for these processes is still unclear.
Fortunately, the genome of C. militaris has recently been sequenced (Zheng et al. 2011a). A total of 9,684 proteincoding genes have been predicted, and 13.7 % of these genes are species-specific, which is significantly higher than the percentage for M. anisopliae (4.8 %) or M. acridum (3.5 %). About 16 % of the C. militaris genes (1,547) are related to pathogen-host interactions. No orthologs of known human mycotoxins have been detected (Zheng et al. 2011a), which is consistent with its safe usage as a medicine. More than 63 % of the total 9,684 genes were expressed during both mycelial growth and fruiting-body
formation. The Zn2Cys6-type transcription factors and MAPK pathway were induced during fruiting, but not the PKA pathway, which differs from the induction of these pathways in other fungi. The complete sequencing of the C. militaris genome will facilitate functional studies of interesting genes and elucidate the genetic background for biosynthesis of bioactive components, insect pathogenicity, and isolate degeneration (Zheng et al. 2011a).
Significant genetic differences between wild-type strains and degenerate strains have been detected (Li et al. 2003, 2007a). However, unlike the significant intraspecific genetic diversity of O. sinensis (Zhang et al. 2009), the genetic distance among C. militaris isolates from different localities is extremely low based on nrDNA ITS sequences (K2P distance value <0.01) (Wang et al. 2008).
Some C. militaris proteins have been purified, and some C. militaris genes have been cloned. The purified proteins include an extracellular trypsin-type serine protease P-1-1 (Hattori et al. 2005), a cytotoxic and antifungal protease CMP (Park et al. 2009), an antifungal peptide cordymin (Wong et al. 2011), a haemagglutinin with anti-proliferative activity towards hepatoma cells (Wong et al. 2009), a lectin (CML) that exhibits haemagglutination activity in mouse and rat erythrocytes (Jung et al. 2007), fibrinolytic enzymes (Kim et al. 2006; Cui et al. 2008; Choi et al. 2011), and superoxide dismutase (Wang et al. 2005a). The cloned genes include a glyceraldehyde-3-phosphate dehydrogenase (GPD) gene (Gong et al. 2009), superoxide dismutase genes (Park et al. 2005; Wang et al. 2005b), and the |3-1,3-glucan synthase catalytic subunit gene (Ujita et al. 2006).
Cultivation of C. militaris
Nutritional requirements for stroma growth and production
Insects
The artificial growth and stroma production of C. militaris has been studied in the laboratory on various insect pupae and larvae, most often on the silkworm Bombyx mori, (Gu and Liang 1987; Liang and Gu 1987; Gu et al. 1988; Gong et al. 1993; Zhou et al. 2000; Chen and Ichida 2002; Li 2002; Pan et al. 2002; Sato and Shimazu 2002b; Zhang et al. 2003; Liu 2004; Wen et al. 2004; Li et al. 2006a; Zheng et al. 2008a, b; Chai et al. 2010; Hong et al. 2010; Mu et al. 2010). Other insects used for artificial stroma production are Antherea pernyi (Gu and Liang 1987; Liang and Gu 1987; Gu et al. 1988; Yuan 1988, 1989; Feng et al. 1990; Wang et al. 2002), Mamestra brassicae (Harada et al. 1995; Sato and Shimazu 2002b), Tenebrio molitor (Sato and Shimazu 2002b; Lin et al. 2005), Ostrinia nubilalis (Liang and Gu 1987), Heliothis virescens, H. zea and Spodoptera.
frugiperda (Sanchez-Pena 1990), Andraca bipunctata (Panigrahi 1995), Philosamia cynthia (Jiang and Xun 1996), Spodoptera litura (Sato and Shimazu 2002b), and Clanis bilineata (Song 2009). Chen and Ichida (2002) documented a higher rate of infection and stroma formation in silkworm pupae than in silkworm larvae. Among three varieties of silkworm (Daeseungjam, Bae-gokjam, and Keumokjam), the Daeseungjam variety was found to be the most suitable for stroma formation of C. militaris (Hong et al. 2010).
Natural organic substrates
Because insects are expensive and not always available (Lin et al. 2006c), and because insects can be difficult to handle and thus prone to microbial contamination, alternative organic substrates have been tested for commercial production of C. militaris stromata. Fortunately, cereals with the addition of some organic substances have proven to be good substitutes for insects. Kobayasi (1941) documented stroma production of C. militaris on rice substrate. Since then, rice has been used as the principal ingredient for growing stromata of C. militaris (Basith and Madelin 1968; Chen and Wu 1990; Liang 1990; Ma and Chen 1991; Sung etal. 1993, 1999; Pen 1995; Sung 1996; Wu et al. 1996; Zhang and Liu 1997; Choi et al. 1999; Li 2002; Zhang 2003; Li et al. 2006d; Lin et al. 2006b; Wen et al. 2008b; Chen et al. 2011b).
The porosity of the fruiting medium affects mycelial growth and fruiting-body yield. Porosity increases with grain size and declines with a higher ratio of water to grain during rice medium preparation. In the absence of interstices, mycelia mostly grow only on the surface of the medium and thereby cannot absorb enough nutrition from the substrate. Interstices permit hyphae to grow inside the medium and to obtain sufficient nutrition from the substrate (Kobayasi 1941). A ratio of rice to water from 1:1 to 1:1.35 or slightly higher has been reported to be optimal for stroma production (Sung et al. 1999, 2002; Lin et al. 2006b; Zheng et al. 2008c; Yue 2010), but the optimal ratio may depend upon the rice cultivar and its glutinous quality. Husked rice (popularly known as brown or unpolished rice) is usually used for cultivation of C. militaris. Maximum fruiting-body yield has been obtained with whole rice grain (Wen et al. 2008b).
Other organic materials used for the production of C. militaris stromata include bean powder, corn grain, corn cobs, cotton seed coats, jowar, millet, sorghum, fragments of sunflower floral disks, and wheat grain (Chen and Wu 1990; Zhang and Liu 1997; Li 2002; Li et al. 2004a; Zhao et al. 2006a; Gao and Wang 2008; Wei and Huang 2009). Rice mixed with silkworm pupae has proven to be superior to other substrates and is now routinely used (Ren 1998; Chen.
et al. 2002; Shrestha et al. 2004a, b, 2005a, b; Sung et al. 2002, 2006a, b; Zhao et al. 2006a; Jin et al. 2009). C. militaris strains require a relatively low level of nitrogen, and excessive nitrogen might suppress differentiation of the fruiting-body (Gao et al. 2000a). This probably explains why yields have been observed to be less on insects than on cereals in the culture. Xie et al. (2009a, b) have also shown that brown rice, malt, and soybean are better sources of nutrition for C. militaris than chemically defined media. Agar media are usually not suitable for stroma production (Basith and Madelin 1968; Yahagi et al. 2004).
Hormones and mineral components of the media
Plant hormones such as 2, 4-D, citric acid triamine, colchicines, and others may enhance stroma production by C. militaris (Li et al. 2004a; Wang et al. 2010b; Xiao et al. 2010). Similarly, mineral salts such as K+, Mg +, and Ca2+ at a concentration of 0.1 g/l may increase fruiting yield (Li et al. 2004a). Some elements may enhance the production of bioactive compounds of C. militaris in culture (Dong et al. 2012).
Duration of stroma production and stromata yield
Commercial production must take into account the duration of stroma production. Stromata are usually produced over a period of 35-70 days (Zhang and Liu 1997; Sung et al. 1999; Yue 2010; Du et al. 2010). Zhang and Liu (1997) reported the production period of 35-45 days on rice but 40-70 days on other substrates such as maize, millet, and rice-tussah. Culture duration, however, depends upon the shape and volume of the culture container and the amount of medium. Stroma production has been quantified in some studies. Wu et al. (1996) obtained 25 g of fresh fruit bodies of C. militaris from 50 g of rice medium, while Zhang and Liu (1997) reported a biological transformation rate of 61 % on rice, 58-59 % on millet and rice-tussah media, and 42 % on maize. Production of 18.0 g of stromata (fresh wt.) has been recently obtained from 20 g of rice (Lin et al. 2006a). Nearly 9 g of dry stromata (equivalent to about 68 g of fresh wt.) was produced from 60 g of brown rice supplemented with 10 g of silkworm pupae (Sung et al. 2006b).
Effect of environment
Whereas hormones govern morphogenetic and developmental changes in plant tissue culture, it is environmental factors that govern the change from the somatic phase to the reproductive phase in fungi. The optimal environmental factors for growth of C. militaris stromata are briefly discussed below.
Temperature
Temperature greatly affects C. militaris stroma production. High temperatures (about 25 °C) lead to maximal mycelial growth, but lower temperatures induce and sustain stroma production, viz. temperatures within the range of 18-22 °C are reported as optimal (Sung et al. 1999, 2002; Gao et al. 2000a; Zhao et al. 2006a; Du et al. 2010; Sato and Shimazu 2002b), although a lower range of 14-17 °C (Du et al. 2007) or a higher temperature of 25 °C (Li et al. 2004a; Yue 2010) have also been reported to be appropriate for stroma production. The maturation period of stromata is shorter at 25 °C than at 20 °C (Sato and Shimazu 2002b).
Light
Light is the most important environmental factor affecting C. militaris stroma production and no stromata are produced in darkness (Sato and Shimazu 2002b). Gao et al. (2000a) obtained stromata under light intensities as high as 4,500 lx, but Sato and Shimazu (2002b) indicated that the upper limit was 1,400 lx; in general, 500-1,000 lx is considered optimal (Sung et al. 1999; Gao et al. 2000a; Sato and Shimazu 2002b; Li et al. 2004a; Zhao et al. 2006a; Du et al. 2010), with a 12-h light/dark cycle of 500-1,000 lx (Sung et al. 2002; Chen et al. 2011b). It has been found that 18 h of light at 200 lx was optimal for pigmentation and primordium initiation, whereas 10 h was suitable for fruiting-body growth and yield (Mu et al. 2010). Longer and wider fruiting-bodies were produced with light intensities of 500-1,000 lx than with an intensity of 100 lx (Hong et al. 2010).
Air exchange and humidity
High air exchange in the culture container favors mycelial growth, primordium formation, and biomass yield (Zhang et al. 2010a). Among materials tested for covering culture bottles containing C. militaris, a hydrophobic fluoropore membrane was the best (Zhang et al. 2010a). A high humidity of 70-90 %, which probably matches the humidity experienced by the fungus in nature, favors stroma production. Low humidity causes the medium to dry too quickly. Especially in dry, indoor environments, a humidifier will be required to maintain sufficient humidity.
Inoculum preparation
For production of C. militaris stromata, media can be efficiently inoculated with a liquid culture of the fungus (Sung 1996). The inoculum in liquid culture consists of conidia, hyphal fragments, and hyphal pellets affected by nutrients, culture duration, mode of culture, etc. The liquid culture.
medium usually contains simple sources of carbon (usually in the form of carbohydrates) and nitrogen (inorganic as well as organic) along with mineral salts. The optimum temperature and pH for liquid culture of C. militaris are 20-25 °C and 6.0-8.0, respectively. While C. militaris my-celia produce yellowish-white to orange pigments on solid media in light (Shrestha et al. 2006), liquid cultures of the fungus usually remain colorless. Pellets in liquid culture increase in size as the culture ages (Han et al. 2009). Pellets are not a suitable form for inoculation purposes for three reasons. First, and most importantly, pellets do not grow as fast as conidial or hyphal suspensions on the fruiting medium. Second, pellets frequently fail to induce stroma primor-dia. Third, inoculum consisting of pellets is difficult to quantify. For large-scale preparation of liquid inoculum, shaking and aeration of the liquid are required for the homogeneous growth of mycelium. The quantity of inoculum added to the fruiting medium depends on the volumes of the culture container and the fruiting medium (Sung et al. 2002; Lin et al. 2006b).
Industrial and commercial development
Although more than 400 Cordyceps s.l. species have been described, only about 36 species have been artificially cultivated for the production of fruiting-bodies (Wang 1995; Sung 1996; Li et al. 2006b). Of those species that have been artificially cultivated, only C. militaris has been commercially cultivated; commercial development has focused on C. militaris because of its excellent pharmaceutical effect and short production period (Li et al. 2006b).
Cordyceps militaris has been cultivated in liquid media for harvesting mycelia and on solid media for induction of fruiting-bodies. While conditions for submerged cultivation of C. militaris inoculum have been optimized (Kim et al. 2003; Liu et al. 2008), large-scale production of C. militaris fruiting-bodies currently uses only solid media consisting of artificial substrates or insects (e.g., the silkworm B. mori). Because cultivation on insects is costly, fruiting-bodies of C. militaris are mainly cultivated on artificial media in which rice is the main component. Substrates used for industrial cultivation of C. militaris in China have recently been reviewed (Wang et al. 2009b). They include media for stock culture, pre-culture spawn, and spawn. For each purpose, the ingredients differ depending on the company (Wang et al. 2009b). Although the cost for cultivation of several insects has recently decreased greatly, fruiting-bodies cultivated on insects (1,000 RMB/kg) are twice as expensive to produce as biomass cultivated in artificial media (500 RMB/kg) (Li et al. 2007b). Compared with the extremely high price of O. sinensis, however, the price of C. militaris is affordable.
In the cultivation of C. militaris fruiting-bodies, four pivotal growth periods are usually identified: mycelial culture, pigment induction, stromata stimulation, and fruiting-body production (Lu et al. 2005). Successful cultivation requires proper control of temperature, humidity, and light (Ren et al. 2009).
Cordyceps militaris cultures have two main uses. First, the fruiting-bodies can be directly consumed as food. C. militaris can be used in stewed chicken, stewed duck, soup, hot pot, tea, and so on. Use of C. militaris in soup is very popular in Southeast Asia, especially in Guangdong, Hong Kong, and Taiwan, China. This use has been shown to be safe if consumption is less than 2.5 g/kg of body weight (Che 2003). Second, C. militaris fruiting-bodies and myce-lia can be used as health products and drugs. In China, many health products contain C. militaris, and these include oral liquids, capsules, wines, vinegars, teas, yogurt, and soy sauce (Wang and Yang 2006). Cultures of C. militaris are also used to produce drugs for maintenance of kidney and lung function, anti-aging, regulation of sleep, and chronic bronchitis (Dai et al. 2007). Currently, more than 30 kinds of C. militaris health products and drugs are available on the market (Huang et al. 2010).
Problems and prospects
Degeneration of isolates
Degeneration of isolates is the main problem in C. militaris cultivation. Degeneration is manifested as reduced growth rate, mycelial density, pigmentation, and fruiting-body yield, and also as changes in fruiting-body shape and size. Degeneration can also be evident in the reduced production of desired compounds from fermented cultures. Degeneration in stroma production is related to the kind of material that was used for isolation. If the isolate was derived from multiple ascospores (multi-spores) or tissue, decline in stroma production occurs soon after one or two subcultures (Shrestha et al. 2004b). Degeneration can be delayed if cultures are obtained from single ascospores (Shrestha et al. 2004a; Sung et al. 2006a).
Compatible pairs of single-ascospore or single-conidium strains should be used to study the effects of biotic and abiotic factors on stroma production. With isolates derived from multiple ascospores, multiple conidia, or tissue, stroma production will vary even when culture conditions are constant (Shrestha et al. 2002, 2004b; Liu et al. 2006). Unfortunately, most publications concerning C. militaris culture do not indicate how the isolate was obtained.
Degeneration in colony pigmentation has been observed in C. militaris strains after several subcultures (Sung et al. 2006a). Lin et al. (2010) observed reduced dehydrogenase
activity and pigment in degenerated strains but no change in mating-type or evidence of dsRNA infection. In addition, the formation of heritable white synnemata has been reported from C. militaris isolates in culture (Sato et al. 1997; Wang et al. 2009a). Formation of white synnemata in C. militaris isolates may provide an opportunity for new strain development as is the case for other cultivated mushrooms such as Flammulina velutipes. A few studies have shown that preservation of C. militaris isolates at 4-10 °C helps to maintain the ability of fruiting-body production for up to 6 months (Sung et al. 2006a; Geng et al. 2009). An increased mutation frequency at the DNA level has been associated with degeneration of C. militaris isolates (Li et al. 2003).
The genes involved in C. militaris degeneration have not been identified. In C. militaris, identification of genes and their function is mostly lacking (Zheng et al. 2011b). So far the glyceraldehyde-3-phosphate dehydrogenase gene, the |3-1,3-glucan synthase catalytic subunit gene, and the superoxide dismutase Cu, Zn-SOD gene are known to be involved in phosphorylation, cell wall constitution, and defense against oxidative damage, respectively (Wang et al. 2005b; Ujita et al. 2006; Gong et al. 2009). Recent research has demonstrated that Agrobacterium tumefa-ciens-mediated transformation is useful to elucidate the function of genes in C. militaris (Zheng et al. 2011b); this method should also help in determining which genes are responsible for degeneration of isolates.
Strain development and large-scale cultivation of C.
militaris
Cordyceps militaris cultivars with desirable properties such as high production of stromata and high cordycepin content have recently been developed (Sun et al. 2009; Du et al. 2010). Du et al. (2010) reported a new cultivar of C. mili-taris with high cordycepin yield (24.98 mg/g of fruiting-body dry wt.). Che et al. (2004) obtained a higher yielding and more stable strain of C. militaris by UV mutagenesis. Recently, regeneration of C. militaris from its protoplasts has been studied (Zhou and Bian 2007; Liu et al. 2009; Zhou and Luo 2009). C. militaris isolates from different regions contain different concentrations of active compounds (Wen et al. 2008a). Thus, superior isolates can be selected for propagation.
Cordyceps militaris is usually cultured in small containers (0.5-1.0 l) because the fungus requires high humidity and moisture, and because small containers such as bottles and trays seem to provide an excellent environment for the growth of primordia and stromata. Reusable light plastic containers can reduce the cost of culture. Substantially increasing production and reducing costs will require the development of bed cultivation or other cultivation methods.
Xiong et al. (2010) considered insect-grown C. militaris to be far superior to cereal-grown C. militaris. Huang et al. (2009), however, reported that fruiting-bodies had higher contents of cordycepin and adenosine when cultivated on rice medium than on silkworm chrysalid or wheat medium. The latter authors also observed that cordycepin and adenosine contents were higher in cultivated fruiting-bodies of C. militaris than in natural O. sinensis fruiting-bodies, and that the contents in cultured mycelium of C. militaris were similar to those in O. sinensis fruiting-bodies. Thus, they proposed that rice-grown C. militaris was the best substitute for O. sinensis (Huang et al. 2009). Brown rice along with malt and soybean has been shown to provide sufficient nutrition for C. militaris (Xie et al. 2009a, b). Wu et al. (2012) compared polysaccharides of C. militaris fruiting-bodies cultivated on rice medium and silkworm pupae, and found that the components and structures of polysaccharides differ in fruiting-bodies cultivated in those two types of media. Similarly, metabolomic studies at regular durations is also useful in order to determine the optimum cultivation period of C. militaris (Choi et al. 2010). There are diverse views, however, regarding the medicinal properties of wild and cultivated C. militaris as well as those grown on various organic substances such as cereals and insects (Li et al. 2004b; Huang et al. 2009; Xiong et al. 2010). Future studies on C. militaris will likely identify new organic substrates that are economical and that support high stroma production and a high concentration of bioactive compounds. The sequencing of its complete genome (Zheng et al. 2011a) will facilitate research on the molecular basis of the biology and medicinal qualities of C. militaris.
The phylogenetic classification has divided the genus Cordyceps into five genera in three families that fit the present concept of the fungal tree of life (Sung et al. 2007; Kepler et al. 2012b). Phylogenetic classification is, however, in a state of flux and has resulted in the frequent transfer of Cordyceps species from one genus to another in different families (Sung et al. 2007; Kepler et al. 2012a). Furthermore, Cordyceps spp. growing on cicadas have been phylo-genetically placed under three different genera (Sung et al. 2007; Sato et al. 2012). Outside the mycological world, the transfer of C. sinensis from Cordyceps to the newly established genus Ophiocordyceps has caused some confusion among researchers in the biochemical and pharmacological communities, because of the belief that O. sinensis (syn. C. sinensis) would be different from Cordyceps spp. at the generic level, though they have adapted to the same level of insect-dependent nutrition. All Cordyceps spp. have similar life cycles and have developed mechanisms to invade insects and grow on them; the major difference being the locality where they grow and the host insect they infect.
Given the diversity of host insects and geographical regions, Cordyceps spp. differ widely in shape, size, color, texture, position of fertile parts, and other micromorphological characters. Among all Cordyceps spp. s.l., O. sinensis has a distinct ecological niche in that it grows at high altitudes in the alpine region. Cordyceps s.l. is a broad genus that encompasses clavicipitaceous fungi that grow on insects as well as on fungi. Cordyceps species that grow on fungi have been phylogenetically placed in the new genera Elaphocor-dyceps and Tyrannocordyceps (Sung et al. 2007; Kepler et al. 2012b). The rest of the Cordyceps species grow on insects and spiders. Cordyceps species remain dormant in soil until they get in contact with a host. They all grow inside the host body and form endosclerotium before emerging from the host body in the summer. Besides their morphological diversity, they also differ widely in cultivability. C. militaris is the most successfully cultivated species (Kobayasi 1941; Sung 1996). As stated above, we believe that C. militaris is worth promoting as a medicinal fungus, being the best described and cultivable Cordyceps species.
Acknowledgments The first author acknowledges the support of the Green Energy Mission/Nepal, Kathmandu, Nepal and a grant from the Next-Generation BioGreen 21 Program (PJ008154 and PJ008321), Rural Development Administration, Republic of Korea, during the preparation of the manuscript. The second author acknowledges the support of the National Natural Science Foundation of China (No. 30870450). The authors also sincerely thank the unknown reviewer for the helpful comments and suggestions.
Alexopoulos CJ, Mims CW, Blackwell M (1996) Introductory mycology, 4th edn. Wiley, New York
Au D, Wang L, Yang D et al (2012) Application of microscopy in authentication of valuable Chinese medicine I—Cordyceps sinensis, its counterfeits, and related products. Microsc Res Tech 75:54-64
Basith M, Madelin MF (1968) Studies on the production of perithecial stromata by Cordyceps militaris in artificial culture. Can J Bot 46:473-480
Cannon PF, Kirk PM (2000) The philosophy and practicalities of amalgamating anamorph and teleomorph concepts. Stud Mycol 45:19-25
Chai JP, Xie DY, Tian XJ et al (2010) Study on defense reactions of silkworm, Bombyx mori to Cordyceps militaris. Southwest China JAgric Sci 23:1308-1313
Chan S, Liu BL, Zhao ZZ et al (2011) Studies on macroscopic and microscopic identification of Cordyceps sinensis and its counterfeits. China J Chin Mater Med 36:1141-1144
Che ZM (2003) Assessment on edible safety of artificially-cultivated Cordyceps militaris fruiting bodies. Edible Fungi 25(3):45-46
Che ZM, Wang Y, Zhou LL et al (2004) Study on the breeding of a new variety of Cordyceps militaris by mutated with ultraviolet radiation. Food Ferment Ind 30(8):35-38
Chen QW (1997) Study on Cordyceps (Fr.) fungi in Shennongjia forest district. Hubei Agric Sci 6:49-52
Chen RY, Ichida M (2002) Infection of the silkworm, Bombyx mori, with Cordyceps militaris. J Insect Biotechnol Sericol 71:61-63 Chen SZ, Wu PJ (1990) A brief introduction to bottle culture technique of Cordyceps militaris. Edible Fungi (04):31 Chen YQ, Piao RZ, Jin YS et al (2002) Study on the artificial good quality and high output cultivation technique of Cordyceps mili-taris. Edible Fungi China 21(5):20-22 Chen CS, Hseu RS, Huang CT (2011a) Quality control of Cordyceps sinensis: teleomorph, anamorph, and its products. In: Shoyama Y (ed) Quality control of herbal medicines and related areas. InTech, Croatia, pp 223-238
Chen YS, Liu BL, Chang YN (2011b) Effects of light and heavy metals on Cordyceps militaris fruit body growth in rice grain-based cultivation. Korean J Chem Eng 28:875-879 Choi IY, Choi JS, Lee WH et al (1999) The condition of production of artificial fruiting body of Cordyceps militaris. Korean J Mycol 27:243-248
Choi JN, Kim JY, Lee MY et al (2010) Metabolomics revealed novel isoflavones and optimal cultivation time of Cordyceps militaris fermentation. J Agric Food Chem 58:4258-4267 Choi D, Cha WS, Park N et al (2011) Purification and characterization of a novel fibrinolytic enzyme from fruiting bodies of Korean Cordyceps militaris. Bioresour Technol 102:3279-3285 Cui JD, Zhang BZ (2011) Comparison of culture methods on exopolysaccharide production in the submerged culture of Cordyceps militaris and process optimization. Lett Appl Microbiol 52:123-128
Cui L, Dong MS, Chen XH et al (2008) A novel fibrinolytic enzyme from Cordyceps militaris, a Chinese traditional medicinal mushroom. World J Microbiol Biotechnol 24:483-489 Dai JJ, Fan T, Wu CH et al (2007) Summarization of the study on the artificial cultivation of Cordyceps militaris Link. J Anhui Agric Sci 35:5469-5471
Das SK, Masuda M, Sakurai A et al (2010) Medicinal uses of the mushroom Cordyceps militaris: current state and prospects. Fito-terapia 81:961-968
de Bary A (1867) Zur Kenntniss insectentoedtender Pilze. Bot Ztg 25:2-28
de Bary A (1887) Comparative morphology and biology of the fungi, mycetozoa and bacteria. Clarendon, Oxford de Cesati V (1861) Appunti per una futura Crittogamologia insubrica.
Comment Soc Crittogam Ital 2:60-75 Ding YH, Li Y, Zhu JH et al (1995) Microscopic studies of Cordyceps militaris under cultivated conditions. Acta Edulis Fungi 2(2):48-52 Dong CH, Yao YJ (2011) On the reliability of fungal materials used in studies on Ophiocordyceps sinensis. J Ind Microbiol Biotechnol 38:1027-1035
Dong JZ, Lei C, Ai XR et al (2012) Selenium enrichment on Cordy-ceps militaris Link and analysis on its main active components. Appl Biochem Biotechnol 166:1215-1224 Du ST, Zhou FL, Chen DY (2007) Studies of temperature influences on Cordyceps militaris growth. J Northwest A F Univ (Nat Sci Ed) 35:159-162
Du AL, Zhang X, Zhang HZ (2010) A new high cordycepin Cordyceps militaris cultivar ‘Haizhou 1’. Acta Hortic Sin 37:1373-1374 Farlow WG, Seymour AB (1888) A provisional host-index of the fungi of the United States: part 1. Polypetalae, Cambridge Feng HL, Guo WC, Zhang RC et al (1990) Histological studies on Cordyceps militaris (L.: Fr.) Link. Acta Mycol Sin 9:1-5 Gams W (1971) Cephalosporium-artige schimmelpilze (Hyphomy-cetes). Gustav Fischer, Stuttgart
Gams W, Hodge KT, Samson RA et al (2005) Proposal to conserve the name Isaria (anamorphic fungi) with a conserved type. Taxon 54:537
Gao XH (2008) Mating system of Cordyceps militaris. Acta Edulis Fungi 15(1):6-10
ao GSY, Wang FZ (2008) Research of commercialized cultivation technology on Cordyceps militaris. North Hortic 9:212-215 Gao XH, Wu W, Qian GC et al (2000a) Study on influences of abiotic factors on fruitbody differentiation of Cordyceps militaris. Acta Agric Shanghai 16(Suppl):93-98
Gao XH, Wu W, Qian GC et al (2000b) Study on relationship between effect of pairing single-ascospore strains of Cordyceps militaris on fruitbody formation and asexual conidiogenous structure. Acta Agric Shanghai 16(Suppl):85-92
Gao Q, Jin K, Ying SH et al (2011) Genome sequencing and comparative transcriptomics of the model entomopathogenic fungi Meta-rhizium anisopliae and M. acridum. PLoS Genet 7:e1001264 Geiser DM, Frisvad JC, Taylor JW (1998) Evolutionary relationships in Aspergillus section Fumigati inferred from partial (5-tubulin and hydrophobin DNA sequences. Mycologia 90:831-845 Geng LJ, He LL, Yu-Yang E et al (2009) Effect of different preservation conditions on mycelial growth and fruit-body yield of Cor-dyceps militaris. J Shenyang Agric Univ 40:165-168 Gong CL, Wu YL, Zhu JH et al (1993) Artificial culture and composition analysis of silkworm Cordyceps militaris. Edible Fungi China 12(4):21-23
Gong CL, Pan ZH, Zheng XJ et al (2006a) Anti-oxidation of cultured Cordyceps militaris growing on silkworm pupa. In: Proceedings of International Workshop on Silk handcrafts cottage industries and silk enterprises development in Africa, Europe, Central Asia and the Near East, & Second Executive Meeting of Black, Caspian seas and Central Asia Silk Association (BACSA), Bursa, Turkey, pp 615-620
Gong CL, Pan ZH, Zheng XJ et al (2006b) Immunoregulation function of artificially Cordyceps militaris growing on pupae of silkworm Bombyx mori for mice. In: Proceedings of International Workshop on Silk handcrafts cottage industries and silk enterprises development in Africa, Europe, Central Asia and the Near East, & Second Executive Meeting of Black, Caspian seas and Central Asia Silk Association (BACSA), Bursa, Turkey, pp 633-638 Gong ZH, Su Y, Huang L et al (2009) Cloning and analysis of glyceraldehyde-3-phosphate dehydrogenase gene from Cordyceps militaris. Afr J Agric Res 4:402-408 Gray GR (1858) Notices of insects that are known to form the bases of fungoid parasites. British Museum, London Gu HS, Liang MY (1987) Study on the manual cultivation of Cordyceps militaris. Pharm Inf Bull 5:51-52 Gu HS, Liang MY, Yuan GH et al (1988) Preliminary study on artificial cultivation of Cordyceps militaris using the pupae of Bombyx mori and Antheraeapernyi. Sci Seric 14:108-110 Gu YX, Wang ZS, Li SX et al (2007) Effect of multiple factors on accumulation of nucleosides and bases in Cordyceps militaris. Food Chem 102:1304-1309
Guo YL, Li CY (2000) Cordyceps kyushuensis new to China. Myco-systema 19:296
Halpern GM (1999) Cordyceps: China’s healing mushroom. Avery, New York
Han YF, Liang JD, Du W et al (2009) Research progress about some problems on Cordyceps militaris. Microbiol China 36:1423-1428 Harada Y, Akiyama N, Yamamoto K et al (1995) Production of Cordyceps militaris fruit body on artificially inoculated pupae of Mames-tra brassicae in the laboratory. Trans Mycol Soc Jpn 36:67-72 Harris SD (2001) Genetic analysis of ascomycete fungi. In: Talbot N (ed) Molecular and cellular biology of filamentous fungi: a practical approach. Oxford University Press, Oxford, pp 47-58 Hattori M, Isomura S, Yokoyama E et al (2005) Extracellular trypsinlike proteases produced by Cordyceps militaris. J Biosci Bioeng 100:631-636
Hawksworth DL (2011) A new dawn for the naming of fungi: impacts of decisions made in Melbourne in July 2011 on the future publication and regulation of fungal names. IMA Fungus 2:155-162
HeLL, Han CL, Fan WLetal (2010) Effects of mineral elements on fruit-body yield and mycelia growth rate of different colony types of Cordyceps militaris in subculturing. Microbiol China37:1331-1340 He SQ, Wang SX, Luo JC et al (2011a) The re-study for morphology of Ophiocordyceps sinensis and Hirsutella sinensis. Microbiol China 38:1730-1738
He Y, Liu LD, Zhao YH et al (2011b) Review on optimum cultivation of Cordyceps militaris. Ludong Univ J (Nat Sci Ed) 27:64-70 Hitchcock SW (1961) Pupal mortality of the orange-striped oakworm. J Econ Entomol 54:962-964
Hodge KT, Gams W, Samson RA et al (2005) Lectotypification and status of Isaria Pers.: Fr. Taxon 54:485-489 Hong IP, Kang PD, Kim KY et al (2010) Fruit body formation on silkworm by Cordyceps militaris. Mycobiology 38:128-132 Hsu TH, Shiao LH, Hsieh C et al (2002) A comparison of the chemical composition and bioactive ingredients of the Chinese medicinal mushroom DongChongXiaCao, its counterfeit and mimic, and fermented mycelium of Cordyceps sinensis. Food Chem 78:463-469
Hu Y, Kang TG, Zhao ZZ (2003) Studies on microscopic identification of animal drugs’ remnant hair (1): identification of Cordyceps sinensis and its counterfeits. Nat Med 57:163-171 Huang SJ, Tsai SY, Lee YL et al (2006) Nonvolatile taste components of fruiting bodies and mycelia of Cordyceps militaris. Food Sci Technol 39:577-583
Huang L, Li QZ, Chen YY et al (2009) Determination and analysis of cordycepin and adenosine in the products of Cordyceps spp. Afr J Microbiol Res 3:957-961
Huang NL, Lin ZB, Chen GL (eds) (2010) Medicinal and edible fungi.
Shanghai Scientific and Technological Literature, Shanghai Hywel-Jones NL (2002) Multiples of eight in Cordyceps ascospores. Mycol Res 106:2-3
Ikeda R, Nishimura M, Sun Y et al (2008) Simple HPLC-UV determination of nucleosides and its application to the authentication of Cordyceps and its allies. Biomed Chromatogr 22:630-636 Jenkins WA (1934) The development of Cordyceps agariciformia. Mycologia 26:220-243
Jiang BL, Xun YG (1996) Studies on culture of Cordyceps militaris on Philosamia cynthia. Nat Plant Resour China 24(2):12-13 Jiang Y, Yao YJ (2002) Names related to Cordyceps sinensis anamorph. Mycotaxon 84:245-254
Jiang Y, Yao YJ (2003) Anamorphic fungi related to Cordyceps sinensis. Mycosystema 22:161-176
Jiang Y, Yao YJ (2005) ITS sequence analysis and ascomatal development of Pseudogymnoascus roseus. Mycotaxon 94:55-73 Jin LY, Du ST, Ma L et al (2009) Optimization on mathematical model of basic medium of Cordyceps militaris cultivation. J Northwest A F Univ (Nat Sci Ed) 37(11):175-179 Jing YB (1987) A summary report on domestication of Cordyceps militaris strain. Edible Fungi 02:5-6 Jones K (1997) Cordyceps: tonic food of ancient China. Sylvan, Washington
Jung EC, Kim KD, Bae CH et al (2007) A mushroom lectin from ascomycete Cordyceps militaris. Biochim Biophys Acta, Gen Subj 1770:833-838
Kamata N (1998) Periodic outbreaks of the beech caterpillar, Quad-ricalcarifera punctatella, and its population dynamics: the role of insect pathogens. In: McManus ML, Liebhold AM (eds) Population dynamics, impacts, and integrated management of forest defoliating insects. USDA Forestry Service, Northeastern Research Station, pp 34-46
Kamata N (2000) Population dynamics of the beech caterpillar, Synty-pistis punctatella, and biotic and abiotic factors. Popul Ecol 42:267-278
Kawamura S (1955) Icones of Japanese fungi. Kazama Shobo, Tokyo 8:821-845.
Kepler RM, Sung GH, Ban S et al (2012a) New teleomorph combinations in the entomopathogenic genus Metacordyceps. Mycologia 104:182-197
Kepler RM, Sung GH, Harada Y et al (2012b) Host jumping onto close relatives and across kingdoms by Tyrannicordyceps (Clavicipitaceae) gen. nov. and Ustilaginoidea (Clavicipitaceae). Am J Bot 99:552-561 Khan MA, Tania M, Zhang DZ et al (2010) Cordyceps mushroom: a potent anticancer nutraceutical. Open Nutraceuticals J 3:179-183 Kim SW, Hwang HJ, Xu CP et al (2003) Optimization of submerged culture process for the production of mycelial biomass and exopolysaccharides by Cordyceps militaris C738. J Appl Microbiol 94:120-126
Kim JS, Sapkota K, Park SE et al (2006) A fibrinolytic enzyme from the medicinal mushroom Cordyceps militaris. J Microbiol 44:622-631
Kirk PM, Cannon PF, David JC et al (eds) (2001) Dictionary of the fungi, 9th edn. CAB International, Oxon Kobayasi Y (1941) The genus Cordyceps and its allies. Sci Rep Tokyo Bunrika Daigaku B 84(5):53-260
Kobayasi Y (1981) Revision of the genus Cordyceps and its allies 1. Bull Natl Sci Mus Tokyo 7:1-13
Kobayasi Y (1982) Keys to the taxa of the genera Cordyceps and Torrubiella. Trans Mycol Soc Jpn 23:329-364 Kothe E (2001) Mating-type genes for classical strain improvement in basidiomycetes. Appl Microbiol Biotechnol 56:602-612 Kryukov VY, Yaroslavtseva ON, Lednev GR et al (2011) Local epizootics caused by teleomorphic cordycipitoid fungi (Ascomycota: Hypocreales) in populations of forest lepidopterans and sawflies ofthe summer-autumn complex in Siberia. Microbiology (Russia) 80:286-295
Kuo HC, Su YL, Yang HL et al (2005) Identification of Chinese medicinal fungus Cordyceps sinensis by PCR-single-stranded conformation polymorphism and phylogenetic relationship. J Agric Food Chem 53:3963-3968
Kuo HC, Su YL, Yang HL et al (2006) Differentiation of Cordyceps sinensis by a PCR-single-stranded conformation polymorphism-based method and characterization of the fermented products in Taiwan. Food Biotechnol 20:161-170 Kwon JS, Lee JS, Shin WC et al (2009) Optimization of culture conditions and medium components for the production of mycelial biomass and exo-polysaccharides with Cordyceps militaris in liquid culture. Biotechnol Bioprocess Eng 14:756-762 Lagerberg T (1922) Smarre meddelanden: Cordiceps militaris (L.)
Link i Sverige. Svensk Bot Tidskr 16:285-290 Leatherdale D (1970) The arthropod hosts of entomogenous fungi in Britain. Entomophaga 15:419-435
Li X (2002) Man made cultivates of Cordyceps militaris (L) Link. J Microbiol (China) 22(6):56-57
Li MN, Wu XJ, Li CY et al (2003) Molecular analysis of degeneration of artificial planted Cordyceps militaris. Mycosystema 22:277-282 Li CB, Tong XD, Bai J et al (2004a) Artificial stromata production of Cordyceps militaris. J Dalian Natl Univ 6(5):29-31 Li SP, Song ZH, Dong TT et al (2004b) Distinction of water-soluble constituents between natural and cultured Cordyceps by capillary electrophoresis. Phytomedicine 11:684-690 Li CR, Huang B, Nam SH et al (2006a) Identification of a strain RCEF0718 with antineoplastic activity. J Laiyang Agric Coll (Nat Sci) 23:263-267
Li CR, Nam SH, Geng DG et al (2006b) Artificial culture of seventeen Cordyceps spp. Mycosystema 25:639-645 Li SP, Yang FQ, Tsimb KWK (2006c) Quality control of Cordyceps sinensis, a valued traditional Chinese medicine. J Pharm Biomed Anal 41:1571-1584
Li SZ, Xia FN, Yang XB (2006d) Comparative studies on the cultivation of 5 selected strains of Cordyceps militaris. Edible Fungi China 25(6):15-16, 19
Li MF, He J, Ding L et al (2007a) Single spore strains without producing fruit body isolated from Cordyceps militaris and their RAPD analysis. Southwest China J Agric Sci 20:547-550 Li S, Zhao YH, Zhu CJ et al (2007b) Development and prospect of Cordyceps militaris. Natl Conf Exch Trans Forest Food Med 2007:55-58
Li J, Chen GS, Fang QM et al (2010) Comparative study on cultivated Cordyceps militaris and wild Cordyceps sinensis. J Chengdu Univ TCM 33(3):82-84
Li CL, Liu XL, Zheng XQ et al (2011) Mutagenesis of Cordyceps militaris protoplast for high production of exopolysaccharide. Ind Microbiol 41(2):51-56
Liang ZQ (1990) Anamorph of Cordyceps militaris and artificial culture of its fruitbody. Southwest China J Agric Sci 3(2):1-6 Liang ZQ (2001) A corroboration of the anamorph of Cordyceps militaris -Paecilomyces militaris Liang sp. nov. Acta Edulis Fungi 8(4):28-32 Liang MY, Gu HS (1987) Success in artificial cultivation of Cordy-ceps. J Shenyang Agric Univ 18:103-104 Liang Y, Zhang GZ, An MP et al (2005) Cordyceps militaris: asco-spore germination and cultural characteristics of progeny population. Mycosystema 24:525-532
Lin QY, Song B, Li TH et al (2005) Studies on Cordyceps militaris (L.: Fr.) Link infecting pupae of Tenebrio molitor L. Mycosystema 24 (Suppl):322-326
Lin QY, Song B, Li TH (2006a) Advances in the studies on Cordyceps militaris. Microbiol China 33(4):154-157 Lin QY, Song B, Zhong YJ et al (2006b) Optimization of some cultivation conditions of Cordyceps militaris. Edible Fungi China 25(6):17-19
Lin QY, Zhong YJ, Li TH et al (2006c) Recent research advances in Cordyceps biology. Acta Edulis Fungi 13(2):93-98 Lin QQ, Qiu XH, Zheng ZL et al (2010) Characteristics of the degenerate strains of Cordyceps militaris. Mycosystema 29:670-677 Liu ZX (2004) Cultivation and the infectious ways to silkworm chrysalis with liquid spawn of Cordyceps militaris. J Huazhong Agric Univ 23:58-60
Liu D, He LL, Wang ZQ et al (2006) The influences of subculture of Cordyceps militaris to colonial morphology and fruit-body yield. J Shenyang Agric Univ 37:538-541 Liu MM, Ning SY, Cui XY et al (2008) Optimization of submerged culture condition for Cordyceps militaris using response surface methodology. Chin Agric Sci Bull 24(5):127-131 Liu XL, Zhou JZ, Huang KH (2009) Study on preparation and regeneration of protoplast of Cordyceps militaris. Acta Agric Jiangxi 21(9):119-120
Liu HJ, Hu HB, Chu C et al (2011) Morphological and microscopic identification studies of Cordyceps and its counterfeits. Acta Pharm Sin B 1:189-195
Lu JM, Zeng ZJ, He HQ (2005) Culture technique of Cordyceps militaris on artificial media. Guangdong Agric Sci 2:88-89 Ma HT, Chen SZ (1991) Status and prospects on artificial cultivation of Cordyceps militaris. J Jinzhou Med Coll 12:63-65 Ma T, Feng Y, Wu XP et al (2007) Primary investigation of a host insect of Cordyceps militaris and analysis of its main ingredients. For Res 20:63-67
Ma LP, Zhao JF, Liu KY et al (2008) Research on the isolation condition of Cordyceps militaris protoplast. J Anhui Agric Sci 36:14612-14613, 14616
Mains EB (1958) North American entomogenous species of Cordy-ceps. Mycologia 50:169-222
McKenna DJ, Jones K, Hughes K (2002) Botanical medicines: the desk reference for major herbal supplements, 2nd edn. Haworth Herbal, New York
Miller JS, Funk VA, Wagner WL et al (2011) Outcomes of the 2011 botanical nomenclature section at the XVIII International Botanical Congress. Phytokeys 5:1-3
Moore RT (1964) Fine structure of mycota: 12 Karyochorisis – somatic nuclear division – in Cordyceps militaris. Z Zellforsch 63:921937
Mu X, Jia CF, Chen S et al (2010) Effects of light hours on growth and development of Cordyceps militaris. J Hebei Agric Sci 14 (12):20-21
Müller-Kögler E (1965) Cordyceps militaris (Fr.) Link: Beobachtungen und Versuche anlässlich eines Fundes auf Tipula paludosa Meig. (Dipt., Tipul.). Z Angew Entomol 55:409-418 Ni H, Li HH, Huang WF et al (2007) Research and product development of Cordyceps militaris and its bioactive substances. Rev Sci Technol 25(15):75-79
Norvell LL (2011) Melbourne approves a new code. Mycotaxon 116:481-490
Pan ZH, Gong CL, Zhu JZ (2002) Technology and application for industrial cultivation of Cordyceps militaris on pupae of Bombyx mori. Jiangsu Seric 24(3):21-24
Panigrahi A (1995) Fungus C. militaris infestation in the pupa of the tea pest Andraca bipunctata Walker. Environ Ecol 13:942-946 Park NS, Lee KS, Sohn HD et al (2005) Molecular cloning, expression, and characterization of the Cu, Zn superoxide dismutase (SOD1) gene from the entomopathogenic fungus Cordyceps militaris. Mycologia 97:130-138
Park BT, Na KH, Jung EC et al (2009) Antifungal and anticancer activities of a protein from the mushroom Cordyceps militaris. Korean J Physiol Pharm 13:49-54
Paterson RRM (2008) Cordyceps—a traditional Chinese medicine and another fungal therapeutic biofactory? Phytochemistry 69:1469-1495 Pen X (1995) The cultivation of Cordyceps militaris fruitbody on artificial media and the determination of SOD. Acta Edulis Fungi 2(3):25-28
Petch T (1942) Notes on entomogenous fungi. Trans Br Mycol Soc 25:250-265
Petch T (1948) A revised list of British entomogenous fungi. Trans Br Mycol Soc 31:286-304
Pettit RH (1895) Studies in artificial cultures of entomogenous fungi.
Cornell Univ Agric Exp Sta Bot Entomol Div Bull 97:339-378 Poggeler S (2001) Mating-type genes for classical strain improvements of ascomycetes. Appl Microbiol Biotechnol 56:589-601 Ren SS (1998) Technical processes for artificial cultivation of Cordyceps militaris with high quality and quantity. Edible Fungi China 17(1):22-23
Ren WY, Zhao H, Wu ZK (2009) Techniques for fast and high yielding cultivation of the valuable edible and medicinal mushroom, Cor-dyceps militaris. China Agric Technol Ext 25(5):28-29 Roumeguere C (1884) Les Spheriacees entomogenes (1). Rev Mycol Toulouse 6:148-154
Sanchez-Pena SR (1990) Some insect and spider pathogenic fungi from Mexico with data on their host ranges. Fla Entomol 73:517-522
Sato H, Shimazu M (2002a) Homothallism in Cordyceps militaris. In: Book of abstracts, 7th International Mycological Congress, Oslo, p 311
Sato H, Shimazu M (2002b) Stromata production for Cordyceps mil-itaris (Clavicipitales: Clavicipitaceae) by injection of hyphal bodies to alternative host insects. Appl Entomol Zool 37:85-92 Sato H, Shimazu M, Kamata N (1994) Detection of Cordyceps milita-ris Link (Clavicipitales: Clavicipitaceae) by burying pupae of Quadricalcarifera punctatella Motschulsky (Lepidoptera: Noto-dontidae). Appl Entomol Zool 29:130-132 Sato H, Kamata N, Shimazu M (1997) Aerial infection of Cordyceps militaris Link (Clavicipitales: Clavicipitaceae) against larvae of Quadricalcarifera punctatella (Motschulsky) (Lepidoptera: Noto-dontidae). Appl Entomol Zool 32:249-252 Sato H, Ban S, Masuya H et al (2012) Reassessment of type specimens of Cordyceps and its allies, described by Dr. Yosio Kobayasi and
preserved in the mycological herbarium of the National Museum of Nature and Science (TNS). Part 3: Cordyceps s. l. on Cicadi-dae. Mycoscience. doi:10.1007/s10267-011-0170-z Shanor L (1936) The production of mature perithecia of Cordyceps militaris (Linn.) Link in laboratory culture. J Elisha Mitchell Sci Soc 52:99-105
Shrestha B, Sung JM (2005) Notes on Cordyceps species collected from central region of Nepal. Mycobiology 33:235-239 Shrestha B, Nam IS, Kim HG et al (2002) Effect of sectors of isolates on fruiting of Cordyceps militaris. Korean Soc Mycol Newsl 14 (1):98
Shrestha B, Kim HK, Sung GH et al (2004a) Bipolar heterothallism, a principal mating system of Cordyceps militaris in vitro. Biotechnol Bioprocess Eng 9:440-446
Shrestha B, Park YJ, Han SK et al (2004b) Instability in in vitro fruiting of Cordyceps militaris. J Mushroom Sci Prod 2:140-144 Shrestha B, Choi SK, Kim HK et al (2005a) Genetic analysis of pigmentation in Cordyceps militaris. Mycobiology 33:125-130 Shrestha B, Han SK, Lee WH et al (2005b) Distribution and in vitro fruiting of Cordyceps militaris in Korea. Mycobiology 33:178181
Shrestha B, Han SK, Yoon KS et al (2005c) Morphological characteristics of conidiogenesis in Cordyceps militaris. Mycobiology 33:69-76
Shrestha B, Lee WH, Han SK et al (2006) Observations on some of the mycelial growth and pigmentation characteristics of Cordyceps militaris isolates. Mycobiology 34:83-91 Shrestha B, Zhang WM, Zhang YJ et al (2010) What is the Chinese caterpillar fungus Ophiocordyceps sinensis (Ophiocordycipita-ceae)? Mycology 1:228-236
Sierpinska A (1998) Towards an integrated management of Dendroli-mus pini L. In: McManus ML, Liebhold AM (eds) Population dynamics, impacts, and integrated management of forest defoliating insects. USDA Forestry Service, Northeastern Research Station, pp 129-142
Song YD (2009) Inoculation experiment of Cordyceps militaris on Clanis bilineata. J Anhui Agric Sci 37:11010-11011 Sopp OJ (1911) Untersuchungen über insekten-vertilgende Pilze bei den letzten Kieferspinnerepidemien in Norwegen. Skrift Vidensk-Selsk I. Mat-Naturv Klasse No. 2 Christiania Stone R (2008) Last stand for the body snatcher of the Himalayas? Science 322:1182
Sun JD, Xiong ST, Wang P (2009) Study on biological and cultivated characters of Cordyceps militaris SN3. J Fungal Res 7(324):148-152 Sung JM (1996) The insects-born fungus of Korea in color. Kyohak Publishing Co. Ltd., Seoul
Sung JM, Shrestha B (2002) In vitro fruiting of Cordyceps mili-taris. In: Book of abstracts, 7th International Mycological Congress, Oslo, p 113
Sung GH, Spatafora JW (2004) Cordyceps cardinalis sp. nov., a new species of Cordyceps with an east Asian-eastern North American distribution. Mycologia 96:658-666 Sung JM, Kim CH, Yang KJ et al (1993) Studies on the distribution and utilization of Cordyceps militaris and C. nutans. Korean J Mycol 21:94-105
Sung JM, Choi YS, Lee HK et al (1999) Production of fruiting body using cultures of entomopathogenic fungal species. Korean J Mycol 27:15-19
Sung JM, Choi YS, Shrestha B et al (2002) Investigation on artificial fruiting of Cordyceps militaris. Korean J Mycol 30:6-10 Sung JM, Park YJ, Lee JO et al (2006a) Effect of preservation periods and subcultures on fruiting body formation of Cordyceps militaris in vitro. Mycobiology 34:196-199
Sung JM, Park YJ, Lee JO et al (2006b) Selection of superior strains of Cordyceps militaris with enhanced fruiting body productivity. Mycobiology 34:131-137
Sung GH, Hywel-Jones NL, Sung JM et al (2007) Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud Mycol 57:5-59
Tulasne LR, Tulasne C (1865) Selecta Fungorum Carpología vol. 3. Paris Museum, Paris
Ujita M, Katsuno Y, Suzuki K et al (2006) Molecular cloning and sequence analysis of the |3-1,3-glucan synthase catalytic subunit gene from a medicinal fungus, Cordyceps militaris. Mycoscience 47:98-105
Ulvinen T (1969) Hyonteisissa elavista Cordyceps-lajeista eraista kuroma-asteisista sienista. Pikkutietoja – Notulae. Ann Entomol Fenn 35:234-237
Varitchak B (1927) Sur le développement des périthèces chez le Cordyceps militaris (Linn.) Link. CR Acad Sci 184:622-625 Wang GD (1995) Ecology, cultivation and application of Cordyceps and Cordyceps sinensis. Scientific and Technical Documents, Beijing Wang JF, Yang CQ (2006) Research survey on artificial cultivation and product development of Cordyceps militaris. Lishizhen Med Mater Med Res 17:268-269
Wang XL, Yao YJ (2011) Host insect species of Ophiocordyceps sinensis: a review. ZooKeys 127:43-59 Wang XQ, Chen CQ, Zhang R (2002) Methodological studies on cultivation of Cordyceps militaris on pupae of Antheraea pernyi. J Anhui Agric Sci 30:965,968
Wang ZS, He ZJ, Li SX et al (2005a) Purification and partial characterization of Cu, Zn containing superoxide dismutase from ento-mogenous fungal species Cordyceps militaris. Enzym Microb Technol 36:862-869
Wang ZS, He ZJ, Shen Q et al (2005b) Purification and partial characterization of recombinant Cu, Zn containing superoxide dismutase of Cordyceps militaris in E. coli. J Chromatogr B 826:114-121 Wang M, Sun MH, Zhang YJ et al (2006) Preliminary studies on microflora of Cordyceps sinensis in Tibet. Edible Fungi China 25(Suppl):6-8
Wang L, Zhang WM, Hu B et al (2008) Genetic variation of Cordyceps militaris and its allies based on phylogenetic analysis of rDNA ITS sequence data. Fungal Divers 31:147-155 Wang HJ, Lv ZL, Zhang BP et al (2009a) Study on variability of Cordyceps militaris. Edible Fungi China 28(5):30-31 Wang YL, Zheng SS, Lv GZ et al (2009b) Current status of investigation on media used for artificial cultivation of Cordyceps militaris in China. Edible Fungi 31(1):1-2
Wang H, Wei J, Lin N et al (2010a) Distribution of mating-type genes in fruiting and non-fruiting forms of Cordyceps militaris. Acta Edulis Fungi 17(4):1-4
Wang HJ, Chu Z, Feng L et al (2010b) A comparative study on effect of two plant growth promoters on the growth of Cordyceps militaris. Lishizhen Med Mater Med Res 21:541-542 Wang XY, Rong YW, Xu L et al (2010c) Electrophoretic karyotype analysis of Paecilomyces militaris, the anamorph of Cordyceps militaris. J Anhui Agric Univ 37:716-719 Webster J, Weber RWS (2007) Introduction to fungi, 3rd edn. Cambridge University Press, New York Wei Q, Huang MQ (2009) Effects of nutrient ingredient in culture medium on the growth of Cordyceps militaris. Beijing Agric 27:36-38 Wen L, Zhang YJ, Zhang TB et al (2004) Studies on culture of Cordyceps militaris on silkworm. Jiangsu Agric Sci 1:91-93 Wen L, Tang YL, Yin QF et al (2005) Assays on nutrient and effective ingredients in different parts of Cordyceps militaris. China J Chin Mater Med 30:659-661
Wen L, Weng L, Zhu MW et al (2008a) Content comparison of active constituent in Cordyceps militaris from different forest regions. Sci Silvae Sin 44(8):149-151
Wen TC, Kang JC, Li GR et al (2008b) Effects of different solid culture condition on fruit body and cordycepin output of Cordyceps militaris. Guizhou Agric Sci 36(4):92-94
Wen TC, Li MF, Kang JC et al (2009) A molecular genetic study on the fruiting-body formation of Cordyceps militaris. Korean Soc Mycol Newsl 21(2):76-95
Wong JH, Wang HX, Ng TB (2009) A haemagglutinin from the medicinal fungus Cordyceps militaris. Biosci Rep 29:321-327 Wong JH, Bg TB, Wang HX et al (2011) Cordymin, an antifungal peptide from the medicinal mushroom Cordyceps militaris. Phytomedicine 18:387-392
Wu YH, Zhu SY, Ding YH et al (1996) Artificial cultivation conditions of Cordyceps militaris and the analysis of its fruitbody components. Acta Edulis Fungi 3(2):59-61 Wu W, Gao XH, Cui XM et al (2000a) Review on studies and applications of Cordyceps militaris. Acta Agric Shanghai 16 (Suppl):99-104
Wu W, Gao XH, Qian GC et al (2000b) Studies on ultrastructure of several typical strains of Cordyceps militaris. Acta Agric Shanghai 16(Suppl):78-84
Wu FY, Yan H, Ma XN et al (2012) Comparison of the structural characterization and biological activity of acidic polysaccharides from Cordyceps militaris cultured with different media. World J Microbiol Biotechnol. doi:10.1007/s11274-012-1005-6 Xiao W, Yang JL, Zhu P et al (2009) Non-support of species complex hypothesis of Cordyceps sinensis by targeted rDNA-ITS sequence analysis. Mycosystema 28:724-730 Xiao ZH, Li ZX, Li JZ et al (2010) Influence of additive on growth and differentiation of Cordyceps militaris (L.) fruitbody. Food Ferment Technol 46(3):60-64
Xie CY, Gu ZX, Fan GJ et al (2009a) Production of cordycepin and mycelia by submerged fermentation of Cordyceps militaris in mixture natural culture. Appl Biochem Biotechnol 158:483-492 Xie CY, Liu GX, Gu ZX et al (2009b) Effects of culture conditions on mycelium biomass and intracellular cordycepin production of Cor-dyceps militaris in natural medium. Ann Microbiol 59:293-299 Xiong CH, Xia YL, Zheng P et al (2010) Developmental stage-specific gene expression profiling for a medicinal fungus Cordyceps mil-itaris. Mycology 1:25-66
Yahagi N, Yahagi R, Takano F et al (2004) Growth of ascoscarps from cultured Cordyceps militaris (L.:Fr.) Fr. and Cordyceps formica-rum Kobayasi in an agar medium. Nippon Kingakukai Kaiho 45:15-19
Yokoyama E, Arakawa M, Yamagishi K et al (2006) Phylogenetic and structural analyses of the mating-type loci in Clavicipitaceae. FEMS Microbiol Lett 264:182-191 Yu HM, Wang BS, Huang SC et al (2006) Comparison of protective effects between cultured Cordyceps militaris and natural Cordy-ceps sinensis against oxidative damage. J Agric Food Chem 54:3132-3138
Yuan GH (1988) Domestication and cultivation of Cordyceps from Jilin Province. Edible Fungi (03):8
Yuan GH (1989) Artificial cultivation of Cordyceps militaris. Life World 1:16-17
Yue C (2010) Optimization on Cordyceps militaris’s cultivating conditions. Food Ind 2:60-61
Yue DC, Yang YP, Wang SF et al (1982) Preliminary study on stroma formation of Cordyceps militaris strain (Briefing). J Chin Med Mater (05):7
Yue GGL, Lau CBS, Fung KP et al (2008) Effects of Cordyceps sinensis, Cordyceps militaris and their isolated compounds on ion transport in Calu-3 human airway epithelial cells. J Ethno-pharmacol 117:92-101
Yun SH, Berbee ML, Yoder OC et al (1999) Evolution of the fungal self-fertile reproductive life style from self-sterile ancestors. Proc Natl Acad Sci USA 96:5592-5597
Zare R, Gams W (2001) A revision of Verticillium section Prostrata: IV. The genera Lecanicillium and Simplicilium gen. nov. Nova Hedwig 73:1-50
Zhang XZ (2003) Biological characteristics and cultivation techniques of Cordyceps militaris C-48. Edible Fungi 25(S1):12 Zhang XK, Liu WX (1997) Experimental studies on planting Cordyceps militaris (L. ex Fr.) Link with different culture materials. Edible Fungi China 16(2):21-22
Zhang J, Song DL, Chen JX (2003) Physiological and biochemical changes of the silkworm, Bombyx mori infected by Cordyceps militaris. Acta Entomol Sin 46:674-678 Zhang YJ, Xu LL, Zhang S et al (2009) Genetic diversity of Ophio-cordyceps sinensis, a medicinal fungus endemic to the Tibetan Plateau: implications for its evolution and conservation. BMC Evol Biol 9:290
Zhang JY, Wu KL, Duan J et al (2010a) Influence of air permeability on growth of Cordyceps militaris. Guangdong Agric Sci 4:45-47 Zhang YJ, Sun BD, Zhang S et al (2010b) Mycobiotal investigation of natural Ophiocordyceps sinensis based on culture-dependent investigation. Mycosystema 29:518-527 Zhang YJ, Zhang S, Wang M et al (2010c) High diversity of the fungal community structure in naturally-occurring Ophiocordyceps sinensis. PLoS One 5:e15570
Zhang S, Zhang YJ, Liu XZ et al (2011) Cloning and analysis of the MAT1-2-1 gene from the traditional Chinese medicinal fungus Ophiocordyceps sinensis. Fungal Biol 115:708-714 Zhang YJ, Bai FR, Zhang S et al (2012a) Determining novel molecular markers in the Chinese caterpillar fungus Ophiocordyceps sinensis by screening a shotgun genomic library. Appl Microbiol Bio-technol. doi:10.1007/s00253-012-4028-x Zhang YJ, Li E, Wang CS et al (2012b) Ophiocordyceps sinensis, the flagship fungus of China: terminology, life strategy and ecology. Mycology 3:2-10
Zhao CY, Li H, Zhang M et al (2006a) Optimization on conditions of artificial cultivation of Cordyceps militaris. J Shenyang Agric Univ 37:209-212
Zhao ZZ, Yuen JPS, Wu JL et al (2006b) A systematic study on confused species of Chinese Materia Medica in the Hong Kong market. Ann Acad Med Singap 35:764-769
Zheng QW, Kang JC (2006) Advances of the research on the artificial cultivation and the pharmacological action of silkworm Cordyceps militaris. J Guizhou Univ (Nat Sci) 23:73-76, 80 Zheng QW, Kang JC, Gao SX et al (2008a) Discussion on the infection of Cordyceps militaris on silkworm in the larval stage. J Anhui Agric Sci 36:3253-3254
Zheng QW, Wang YY, Gao SX (2008b) The study of Cordyceps militaris infecting the 5th instar silkworm. Edible Fungi 30(5):32-34 Zheng SS, Xu C, Han NN et al (2008c) Optimum carbon source screening of Cordyceps militaris medium in artificial condition. J Heilongjiang August First Land Reclam Univ 20(1):8-11 Zheng P, Xia YL, Xiao GH et al (2011a) Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol 12:R116 Zheng ZL, Huang CH, Cao L et al (2011b) Agrobacterium tumefaciens-mediated transformation as a tool for insertional mutagenesis in medicinal fungus Cordyceps militaris. Fungal Biol 115:265-274 Zhong S, Ji DF, Chen S et al (2006) Research advancement of Cordyceps militaris. Bull Seric 37(4):6-10 Zhong S, Pan HJ, Fan LF et al (2009) Advances in research of polysaccharides in Cordyceps species. Food Technol Biotechnol 47:304-312
Zhou HY, Bian YB (2007) Identification and selection of high cordycepin-yielding protoplast fusion products of Cordyceps mil-itaris. Acta Edulis Fungi 14(2):65-70 Zhou LH, Luo LM (2009) Preparation and regeneration of protoplasts from Cordyceps militaris. Hubei Agric Sci 48:1621-1624 Zhou GQ, Han DH, Wan XX et al (2000) Behavior of Cordyceps militaris on the tussah and content of polysaccharide in fermentation liquid. J Dalian Inst Light Ind 19(2):108-111 Zhou LH, Wang SM, Luo LM (2009a) The mutagenesis and screening of cordycepin-over-producing strain of Cordyceps militaris by UV treatment. Edible Fungi 31(4):20-21 Zhou XW, Gong ZG, Su Y et al (2009b) Cordyceps fungi: natural products, pharmacological functions and developmental products. J Pharm Pharmacol 61:279-291.
Viện Nghiên cứu Sinh học Ứng dụng
Địa chỉ: Số 39, Ngõ 189/61, Hoàng Hoa Thám, Ngọc Hà, Quận Ba Đình, TP Hà Nội
ĐT: (+84) 2422 118 008 – (+84)962 567 869
Website: https://vbio.vn/
Email: vbiovn1@gmail.com

Giao hàng toàn quốc
-
 Nhận hàng & thanh toán tại nhà
Nhận hàng & thanh toán tại nhà
-
 Miễn phí vận chuyển khi mua 10 PS
Miễn phí vận chuyển khi mua 10 PS
TIN LIÊN QUAN

EDIBLE AND MEDICINAL MUSHROOMS
Ganoderma Karst, belongs to the groups Eumycota, Basidiomycotina, Hymenomycetes, Aphyllophorales, Ganodermatacese, and is a genus of wood decaying polypore fungi of economic importance. Up to now, more...
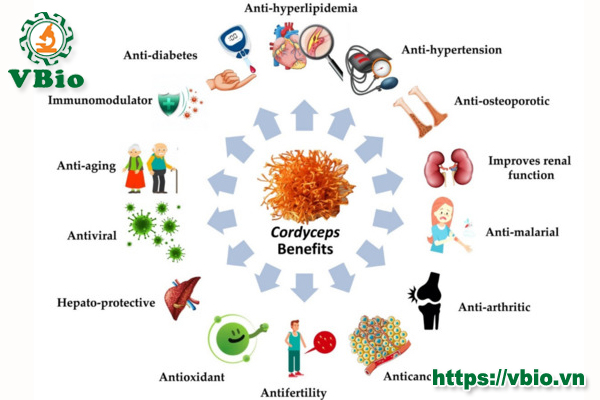
Cordycepin for Health and Wellbeing
Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic Medicinal Fungus Cordyceps with Its Nutraceutical and Therapeutic Potential Abstract: Cordyceps is a rare naturally...
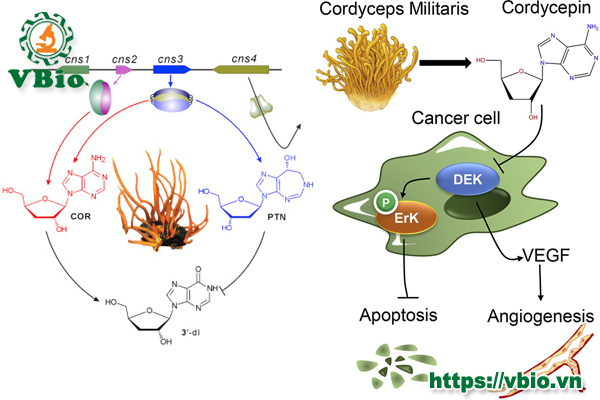
Determination and analysis of cordycepin and adenosine
Determination and analysis of cordycepin and adenosine in the products of Cordyceps spp Cordyceps sinensisis, a kind of precious natural crude drugs and edible mushrooms, were used...
Cordycepin: A Cordyceps Metabolite with Promising Therapeutic Potential
For thousands of years, natural products from medicinal mushroom are being used for the cure of different lethal diseases. Among the huge category of medicinal herbs, the...








 0
0

